
2016 , Vol. 10 >Issue 06: 326 - 332
DOI: https://doi.org/10.3877/cma.j.issn.1674-0807.2016.06.002
E6-AP 基因在乳腺癌MDA-MB-231 细胞中调节膜联蛋白A2 的表达
Copy editor: 刘军兰
收稿日期: 2015-11-10
网络出版日期: 2024-12-04
基金资助
国家自然科学基金资助项目(81172496)四川省教育厅重点项目(13ZA0228)四川省科技厅科技创新苗子工程项目(2016060)
版权
E6-AP gene regulates Annexin A2 expression in breast cancer MDA-MB-231 cells
Received date: 2015-11-10
Online published: 2024-12-04
Copyright
目的
检测E6-AP 基因及膜联蛋白A2(Annexin A2)在乳腺癌MDA-MB-231 细胞中的表达,探讨其对癌细胞增殖、凋亡及浸润转移的影响。
方法
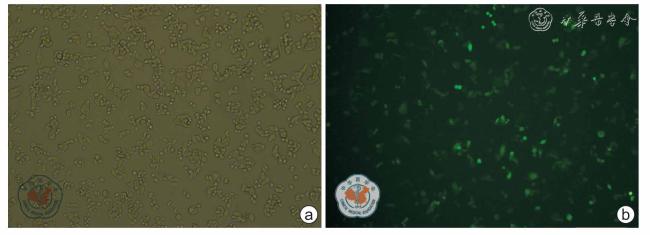
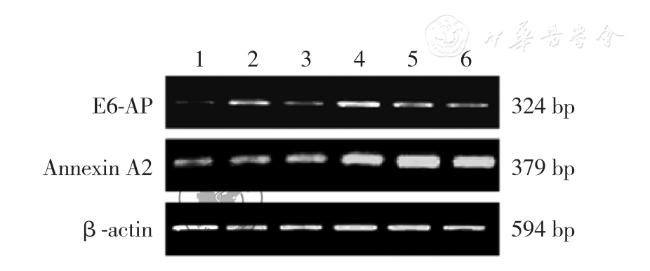
设计1 条无关序列Negative-siRNA 作为阴性对照组和针对E6-AP 基因的3 条特异性E6-AP-siRNAs 片段转染至MDA-MB-231 细胞内作为实验组,未经处理细胞作为空白对照组,加入脂质体处理的细胞为脂质体组,利用RT-PCR 检测干扰E6-AP 后在MDA-MB-231 细胞中E6-AP 和Annexin A2 mRNA 相对表达水平。 选择出转染效率最高的E6-APsiRNA1 组及阴性对照组、空白对照组继续行后续实验。 Western blot 检测干扰E6-AP 后E6-AP 和Annexin A2 在MDA-MB-231 细胞中的蛋白的相对表达水平。 利用CCK-8 试剂盒法、流式细胞术、Transwell 小室侵袭实验分别检测干扰E6-AP 后MDA-MB-231 细胞的增殖、凋亡、侵袭能力。 基因的mRNA 及蛋白表达水平、细胞凋亡率及细胞数的组间比较采用方差分析, 两两比较采用LSD 法,吸光度比较采用重复测量的方差分析。
结果
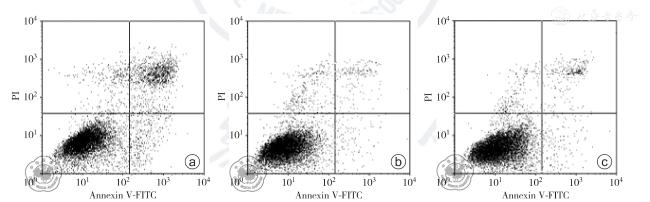
转染72 h 后,E6-AP 基因干扰后各实验组(E6-AP-siRNA1 组、E6-AP-siRNA2 组、E6-AP-siRNA3 组)及空白对照组、阴性对照组及脂质体组中的E6-AP mRNA 相对表达水平分别为0.159±0.003、0.325±0.006、0.229±0.007、0.593 ±0.031、0.594±0.012、0.612±0.016,Annexin A2 mRNA 相对表达水平分别为0.929±0.017、1.013±0.082、0.992±0.024、1.341±0.037、1.323±0.010、1.326±0.012,差异均有统计学意义(F=850.792、417.447,P 均<0.050)。 转染72 h 后,E6-APsiRNA1 组、空白对照组和阴性对照组中E6-AP 及Annexin A2 蛋白相对表达水平为分别为0.271±0.017、0.492±0.018、0.477±0.016 及0.447±0.034、0.887±0.022、0.849±0.033,组间差异均有统计学意义(F=256.850、350.149,P 均<0.050)。 转染24、48、72、96 h 后,E6-AP-siRNA1 组、阴性对照组和空白对照组间比较,不同时间点之间比较,细胞吸光度差异均有统计学意义(F=524.828, P<0.001;F=904.079,P<0.001);分组与时间点存在交互作用(F=28.116, P<0.001)。 转染72 h 后,空白对照组、阴性对照组、E6-AP-siRNA1 组的凋亡率分别为2.959±0.117、3.097±0.070、10.812±0.199,组间差异有统计学意义(F=3110.005,P<0.050)。 Transwell 检测E6-AP-siRNA1 组、空白对照组、阴性对照组中细胞穿透Matrigel 胶到达Transwell 下室的细胞数分别为99±5、96±6、62±7,组间差异有统计学意义(F=55.404,P<0.001)。
结论
干扰E6-AP 基因可使Annexin A2 表达下调,同时可诱导MDA-MB-231 细胞的凋亡,其增殖、侵袭能力也受到抑制。
侯令密 , 谢少利 , 陈茂山 , 李敬东 , Emmanuel Ajedichiga Aduah , 王明浩 , 邓世山 , 幸天勇 , 赵小波 . E6-AP 基因在乳腺癌MDA-MB-231 细胞中调节膜联蛋白A2 的表达[J]. 中华乳腺病杂志(电子版), 2016 , 10(06) : 326 -332 . DOI: 10.3877/cma.j.issn.1674-0807.2016.06.002
Objective
To study the expressions of E6-AP gene and Annexin A2 in breast cancer MDA-MB-231 cells, and explore their effects on the proliferation, apoptosis and invasion and metastasis of cancer cells.
Methods
Negative-siRNA was transfected into MDA-MB-231 cells as negative control,3 designed E6-AP-siRNA sequences were transfected into MDA-MB-231 cells as experimental groups, the untreated cells served as blank control and the cells with liposome treatment served as liposome group. The mRNA expressions of E6-AP and Annexin A2 in MDA-MB-231 cells were detected by RT-PCR after E6-AP interference. The E6-AP-siRNA1 group with the highest transfection efficiency, along with negative control group and blank control group, was selected for the following experiments. The protein expressions of E6-AP and Annexin A2 in MDA-MB-231 cells after E6-AP interference were detected by Western blot. The proliferation, apoptosis and invasion of MDA-MB-231 cells were detected by CCK-8 kit, flow cytometry and Transwell assay respectively after E6-AP interference. Comparison between groups was performed using analysis of variance, pairwise comparison using LSD method. The optical density was compared by repeated measurement analysis of variance.
Results
At 72 h after transfection, E6-AP mRNA expressions in E6-APsiRNA1 group, E6-AP-siRNA2 group, E6-AP-siRNA3 group, blank control group, negative control group and liposome group were 0.159±0.003, 0.325±0.006, 0.229±0.007, 0.593±0.031, 0.594±0.012, 0.612±0.016 respectively, Annexin A2 mRNA expressions were 0.929±0.017, 1.013±0.082, 0.992±0.024,1.341±0.037,1.323±0.010, 1.326±0.012 respectively, indicating statistically significant differences (F=850.792,417.447, both P<0.050). E6-AP and Annexin A2 protein expressions in E6-AP-siRNA1 group,blank control group and negative control group were 0.271±0.017,0.492±0.018,0.477±0.016 and 0.447±0.034,0.887±0.022, 0.849 ± 0.033, respectively, indicating statistically significant differences (F =256.850,350.149, both P <0.050). There was a significant difference in optical density among E6-APsiRNA1 group, negative control group and blank control group at 24,48,72,96 h after the transfection (F=524.828, P<0.001), and there was also a significant difference in optical density among the different time points (F= 904.079, P<0.001). There was an interaction between the grouping and the time points (F=28.116, P<0.001). The apoptosis rates in blank control group, negative control group and E6-AP-siRNA1 group were 2.959±0.117, 3.097±0.070 and 10.812±0.199 respectively, and the difference was significant among groups (F=3110.005, P<0.050). The number of cells which went through Matrigel gel into lower Transwell chamber in E6-AP-siRNA1 group, blank control group and negative control group was 99±5,96±6,62±7,respectively,suggesting a significant difference among groups (F= 55.404,P<0.001).
Conclusion
Interference with E6-AP gene may down-regulate the expression of Annexin A2, induce the apoptosis of MDAMB-231 cells, inhibit the proliferation and invasion ability of MDA-MB-231 cells.
Key words: Breast neoplasms; Cell proliferation; Apoptosis; Neoplasm invasiveness
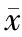 ±s表示,各组间差异比较采用方差分析, 两两比较采用LSD 法,细胞吸光度值比较采用重复测量的方差分析。 P<0.050 为差异具有统计学意义。
±s表示,各组间差异比较采用方差分析, 两两比较采用LSD 法,细胞吸光度值比较采用重复测量的方差分析。 P<0.050 为差异具有统计学意义。 吸光度值的比较( |
| 组别 | 转染时间 | |||
|---|---|---|---|---|
| 24h | 48h | 72h | 96h | |
| 空白对照组 | 0.448±0.027 | 0.494±0.013 | 0.607±0.024 | 0.717±0.020 |
| 阴性对照组 | 0.476±0.044 | 0.496±0.020 | 0.602±0.009 | 0.704±0.020 |
| E6-AP-siRNA1组 | 0.390±0.028 | 0.434±0.026 | 0.480±0.029 | 0.544±0.016 |
| [1] |
Helchowski CM, Skow LF, Roberts KH, et al. A small ubiquitin binding domain inhibits ubiquitin-dependent protein recruitment to DNA repair foci[J]. Cell Cycle,2013,12(24):3749-3758.
|
| [2] |
Salomè M, Campos J,Keeshan K. TRIB2 and the ubiquitin proteasome system in cancer[J].Biochem Soc Trans,2015,43(5):1089-1094.
|
| [3] |
Ando H, Wen ZM, Kim HY, et al. Intracellular composition of fatty acid affects the processing and function of tyrosinase through the ubiquitin-proteasome pathway[J]. Biochem J, 2006,394(Pt 1):43-50.
|
| [4] |
Mattern MR, Wu J, Nicholson B. Ubiquitin-based anticancer therapy:carpet bombing with proteasome inhibitors vs surgical strikes with E1,E2, E3, or DUB inhibitors[J]. Biochim Biophys Acta,2012,1823(11):2014-2021.
|
| [5] |
Hillemanns P,Jentschke M,Evans TG,et al. Detection of E6-AP as a potential therapeutic target in cervical specimen of HPV-infected women[J]. Arch Gynecol Obstet,2014,289(6):1281-1286.
|
| [6] |
Deng S, Jing B, Xing T, et al. Overexpression of annexin A2 is associated with abnormal ubiquitination in breast cancer[J]. Genomics Proteomics Bioinformatics,2012,10(3):153-157.
|
| [7] |
Deng S, Wang J, Hou L, et al. Annexin A1, A2, A4 and A5 play important roles in breast cancer, pancreatic cancer and laryngeal carcinoma, alone and/or synergistically[J]. Oncol Lett,2013,5(1):107-112.
|
| [8] |
Deng S, Huang C. E3 ubiquitin ligases in regulating stress fiber,lamellipodium, and focal adhesion dynamics[J]. Cell Adh Migr,2014,8(1):49-54.
|
| [9] |
Zhang J, Song R, Li Y, et al. Integration of microarray profiles associated with cardiomyopathy and the potential role of E6-AP in apoptosis[J]. Mol Med Rep,2014,9(2):621-625.
|
| [10] |
Lokman NA, Ween MP, Oehler MK, et al. The role of Annexin A2 in tumorigenesis and cancer progression[J]. Cancer Microenviron,2011,4(2):199-208.
|
| [11] |
Li PG, Yang YL,Ge YT. Roles of annexin A2 for the regulation of the growth and cytoskeleton of tumor cells[J]. Sheng Li Ke Xue Jin Zhan,2010,41(6):457-460.
|
| [12] |
Hajjar KA. The biology of annexin A2: from vascular fibrinolysis to innate immunity[J]. Trans Am Clin Climatol Assoc, 2015,126:144-155.
|
| [13] |
Lauritzen SP, Boye TL, Nylandsted J. Annexins are instrumental for efficient plasma membrane repair in cancer cells[J]. Semin Cell Dev Biol,2015,45:32-38.
|
| [14] |
Jeon YR, Kim SY, Lee EJ, et al. Identification of annexin II as a novel secretory biomarker for breast cancer[J]. Proteomics, 2013,13(21):3145-3156.
|
| [15] |
Deng S, Jing B, Xing T, et al. Overexpression of annexin A2 is associated with abnormal ubiquitination in breast cancer [J]. Genomics Proteomics Bioinformatics,2012,10(3):153-157.
|
| [16] |
Wolyniec K, Shortt J, de Stanchina E, et al. E6AP ubiquitin ligase regulates PML-induced senescence in Myc-driven lymphomagenesis[J].Blood,2012,120(4):822-832.
|
| [17] |
Lochab S, Pal P, Kanaujiya JK, et al. Proteomic identification of E6AP as a molecular target of tamoxifen in MCF7 cells [ J].Proteomics,2012,12(9):1363-1377.
|
| [18] |
侯令密, 谢少利, 陈茂山, 等. E6 相关蛋白调节膜联蛋白A2 的表达对三阴性乳腺癌裸鼠移植瘤的影响[J/CD].中华妇幼临床医学杂志:电子版,2016,12(30):274-279.
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}