
2024 , Vol. 18 >Issue 04: 206 - 211
DOI: https://doi.org/10.3877/cma.j.issn.1674-0807.2024.04.003
NF2/YAP信号通路通过FSP1诱导CD24高表达的三阴性乳腺癌细胞铁死亡
收稿日期: 2023-11-14
网络出版日期: 2024-08-20
基金资助
四川省科技厅自然科学基金面上项目(2022NSFSC0775)
四川省基层卫生事业发展研究中心资助项目(SWFZ22-C-82)
南充市科技局市校科技战略合作项目(20SXQT0052)
川北医学院校级科研发展计划项目(CBY22-QNA11)
版权
NF2/YAP signaling pathway induces ferroptosis in triple negative breast cancer cells with high expression of CD24 by FSP1
Received date: 2023-11-14
Online published: 2024-08-20
Copyright
研究NF2和YAP表达对CD24高表达的三阴性乳腺癌细胞铁死亡的影响,探讨铁死亡的相关分子机制。
培养CD24high和CD24lowMDA-MB-231细胞,通过台盼蓝染色(TBA)实验检测经半胱氨酸饥饿、铁死亡诱导剂Erastin和RSL3处理后细胞的死亡情况,TBA法检测丙二醛(MDA)含量,分析细胞铁死亡水平。使用Western blot检测NF2和YAP在CD24high和CD24lowMDA-MB-231细胞中的蛋白表达情况,并用慢病毒分别进行敲除和过表达,观察细胞铁死亡水平,并采用实时定量RT-PCR和Western blot检测成纤维细胞特异蛋白1(FSP1)的表达变化。
经半胱氨酸饥饿、铁死亡诱导剂Erastin和RSL3分别处理后MDA-MB-231细胞死亡率增加,CD24low细胞死亡率高于CD24high细胞(t=14.548,P<0.001;t=8.310,P=0.001;t=8.600,P=0.001)。同时,TBA法结果显示,在含或不含半胱氨酸条件下,CD24high和CD24lowMDA-MB-231细胞的MDA含量差异有统计学意义(t=-3.920,P=0.017;t=11.566,P<0.001);与对照组(DMSO)相比,CD24high和CD24low细胞经铁死亡诱导剂Erastin和RSL3处理后MDA水平增加,其中CD24low细胞的MDA含量比CD24high更高(t=10.763,P=0.006;t=24.067,P<0.001)。Western blot实验结果表明,NF2与YAP在CD24low和CD24high细胞的蛋白含量差异有统计学意义(t=-4.331,P=0.012;t=4.219,P=0.013),CD24low细胞中YAP的蛋白表达更高(t=4.219,P=0.013),而CD24high细胞中NF2的蛋白表达更高(t=-4.331,P=0.012)。在CD24highMDA-MB-231细胞中构建NF2敲除或(和)YAP过表达细胞株,并用实时定量RT-PCR和Western blot检测,结果显示铁死亡关键蛋白FSP1在4组间的表达差异有统计学意义(F=30.297,P<0.001)。同时,检测NF2敲除或(和)YAP过表达后CD24highMDA-MB-231细胞的铁死亡情况,结果表明敲除NF2或(和)过表达YAP后加入铁死亡诱导剂Erastin均可在一定程度上促进细胞死亡(F=38.911,P<0.050),其中在NF2敲除联合YAP过表达条件下,CD24highMDA-MB-231细胞死亡率最高,而在Erastin的基础上添加铁死亡抑制剂Fer-1后细胞的死亡率下降,4组间比较差异无统计学意义(F=0.256,P=0.855)。
在CD24高表达的三阴性乳腺癌细胞中敲除NF2联合过表达YAP会抑制FSP1表达,促进细胞铁死亡。
关键词: 乳腺肿瘤; 铁死亡; 成纤维细胞特异蛋白1; NF2; YAP
蒲卢兰 , 李静佳 , 陈宇 , 周瑜清 , 荣欣欣 , 侯令密 , 周方方 . NF2/YAP信号通路通过FSP1诱导CD24高表达的三阴性乳腺癌细胞铁死亡[J]. 中华乳腺病杂志(电子版), 2024 , 18(04) : 206 -211 . DOI: 10.3877/cma.j.issn.1674-0807.2024.04.003
To observe the impact of NF2 and YAP expression on ferroptosis of triple negative breast cancer cells with high expression of CD24 and explore the related molecular mechanism.
MDA-MB-231 breast cancer cells with high expression of CD24 (CD24high) and low expression of CD24 (CD24low) were cultured. The cells were treated with ferroptosis agonists Erastin, RSL3 and cysteine starvation. The cell death was detected by Trypan blue staining (TBA). Malondialdehyde(MDA)content was measured using the TBA method to analyze the level of cellular ferroptosis. Western blot analysis was used to detect the protein expression of NF2 and YAP in CD24high and CD24low MDA-MB-231 cells. Lentiviral vectors were used to knock out and over-express NF2 and YAP, and changes in cellular ferroptosis were observed. The real-time quantitative RT-PCR and Western blot analysis were used to detect the expression of fibroblast-specific protein 1(FSP1)after NF2 knock out and YAP over-expression.
After treatment with cysteine starvation, the ferroptosis inducers Erastin and RSL3, the death rate of MDA-MB-231 cells increased, with the death rate of CD24low cells significantly higher than that of CD24high cells (t=14.548, P<0.001; t=8.310, P=0.001; t=8.600, P=0.001). Meanwhile, the TBA method results showed that there was a significant difference in MDA content between CD24high and CD24low MDA-MB-231 cells under conditions with or without cysteine (t=-3.920, P=0.017; t=11.566, P<0.001); compared with the control group with DMSO, the MDA levels of CD24high and CD24low cells significantly increased after treatment with the ferroptosis inducers Erastin and RSL3, with the MDA content of CD24low cells significantly higher than that of CD24high cells (t=10.763, P=0.006; t=24.067, P<0.001). Western blot experiments showed that there was a significant difference in the protein content of NF2 and YAP between CD24low and CD24high MDA-MB-231 cells (t=-4.331, P=0.012; t=4.219, P=0.013), with higher expression of YAP protein in CD24low cells (t=4.219, P=0.013) and higher expression of NF2 protein in CD24high cells (t=-4.331, P=0.012). NF2 knockout or (and) YAP overexpression cell lines were constructed in CD24high cells, and real-time quantitative RT-PCR and Western blot analysis were used to detect the expression of the ferroptosis key protein FSP1 among the four groups (F=30.297, P<0.001). Meanwhile, the ferroptosis situation of CD24high MDA-MB-231 cells after NF2 knockout or (and) YAP overexpression was detected, and the results showed that knocking out NF2 or (and) overexpressing YAP could promote cell death to a certain extent after adding the ferroptosis inducer Erastin (F=38.911, P<0.050), with the highest death rate of CD24high MDA-MB-231 cells under the condition of NF2 knockout combined with YAP overexpression. The addition of the ferroptosis inhibitor Fer-1 after Erastin led to a decrease in the death rate of cells, with no statistically significant difference among the four groups (F=0.256, P=0.855).
Knocking out NF2 and overexpressing YAP in CD24high triple negative breast cancer cells can inhibit the expression of FSP1 and promote cell ferroptosis.
Key words: Breast neoplasms; Ferroptosis; Fibroblast-specific protein 1; NF2; YAP
表1 实时荧光定量反转录PCR引物序列 |
| 引物名称 | 引物序列(5’→3’) |
|---|---|
| NF2 | 正向:GTGCCATACTAAGCAGCTTGC |
| 反向:ACACTACAGCTCGGAAATCCA | |
| YAP | 正向:GTCGGAACCCTTCCCTCATTA |
| 反向:GGCCTTCACTGACAGGTAGTA | |
| FSP1 | 正向:ATCGTCCGAGATGATGTTGATGGG |
| 反向:ATTACTGCTACTCCGTACACCACTG | |
| GAPDH | 正向:TGACTTCAACAGCGACACCCA |
| 反向:CACCCTGTTGCTGTAGCCAAA |
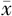 ±s表示,独立样本或组间比较采用t检验,多组间比较采用单因素方差分析(两两比较采用LSD校正)。双侧检验,以P<0.050为差异有统计学意义。
±s表示,独立样本或组间比较采用t检验,多组间比较采用单因素方差分析(两两比较采用LSD校正)。双侧检验,以P<0.050为差异有统计学意义。表2 半胱氨酸饥饿处理后CD24low与CD24high细胞死亡率比较 |
| 组别 | 实验次数 | 含半胱氨酸 | 半胱氨酸饥饿 |
|---|---|---|---|
| CD24low | 3 | 4.60±0.63 | 28.93±0.40 |
| CD24high | 3 | 3.83±0.81 | 12.60±1.82 |
| t值 | 1.301 | 14.548 | |
| P值 | 0.263 | <0.001 |
表3 铁死亡诱导剂处理CD24low与CD24high细胞死亡率比较 |
| 组别 | 实验次数 | DMSO | Erastin | RSL3 |
|---|---|---|---|---|
| CD24low | 3 | 4.93±1.36 | 38.15±2.92 | 39.51±2.16 |
| CD24high | 3 | 3.61±1.12 | 14.73±3.91 | 18.90±1.79 |
| t值 | 1.292 | 8.310 | 8.600 | |
| P值 | 0.266 | 0.001 | 0.001 |
表4 半胱氨酸饥饿处理后CD24low与CD24high细胞MDA水平比较 |
| 组别 | 实验次数 | 含半胱氨酸 | 半胱氨酸饥饿 |
|---|---|---|---|
| CD24low | 3 | 0.32±0.02 | 1.63±0.06 |
| CD24high | 3 | 0.37±0.01 | 0.96±0.08 |
| t值 | -3.920 | 11.566 | |
| P值 | 0.017 | <0.001 |
表5 铁死亡诱导剂处理后CD24low与CD24high细胞MDA水平比较 |
| 组别 | 实验次数 | DMSO | Erastin | RSL3 |
|---|---|---|---|---|
| CD24low | 3 | 0.42±0.04 | 1.76±0.02 | 1.67±0.02 |
| CD24high | 3 | 0.40±0.03 | 1.28±0.08 | 1.22±0.03 |
| t值 | 0.970 | 10.763 | 24.067 | |
| P值 | 0.387 | 0.006 | <0.001 |
表6 NF敲除联合YAP过表达CD24high细胞加药处理后死亡率比较 |
| 组别 | 实验次数 | DMSO | Erastin | Erastin+Fer-1 |
|---|---|---|---|---|
| 对照 | 3 | 5.74±0.50 | 19.89±0.68 | 6.37±2.50 |
| YAP-OV | 3 | 6.64±1.33 | 29.43±0.53 | 6.68±0.78 |
| NF2-KO | 3 | 5.86±1.34 | 36.31±6.10 | 7.42±1.48 |
| YAP-OV+NF2-KO | 3 | 6.19±0.60 | 46.28±0.57 | 7.01±0.71 |
| F值 | 0.469 | 38.911 | 0.256 | |
| P值 | 0.712 | <0.001 | 0.855 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}